







































We’re often told that the distribution of traits among organisms fits neatly with a nested hierarchy or an evolutionary “tree,” thereby supposedly demonstrating common descent. This claim is true, except for when it isn’t. There are numerous cases where it isn’t, and common ancestry cannot explain the distribution of a given trait. (We’ve covered many times how the evidence doesn’t fit with the tree-like expectations of common ancestry, for example here, here, or here.) As the esteemed Cambridge paleontologist and evolutionary biologist Simon Conway Morris (not an ID proponent) has written, “Convergence is ubiquitous.”
Now, a new paper in PLOS Biology would seem to agree, as it opens by stating, “Convergent evolution, the repeated evolution of similar phenotypes, is widespread in nature…” The title of the paper is “Genetic parallelism underpins convergent mimicry coloration in Lepidoptera across 120 million years of evolution,” and it recounts newly discovered evidence of supposed convergent evolution — not just in their bodies, but even at the genetic level no less — in certain flying insects. An article at Science Daily explains what they investigators found:
Researchers found that distantly related butterflies and moths have reused the same pair of genes for over 120 million years to produce strikingly similar warning colors. Rather than altering the genes themselves, evolution modifies how they’re switched on and off. This discovery hints that life may evolve in more predictable ways than previously believed.
It continues:
evolution acted on regulatory elements, often described as genetic “switches,” that control when and where these genes are activated. In butterflies, these switches were modified in similar ways across species. In the moth, scientists found a surprising twist. It used an inversion mechanism (a large chunk of DNA flipped backwards) that closely mirrors a strategy seen in one of the butterfly species.
The catch is that these two species are distantly related, so it’s not thought that the common ancestor would have had this genetic trait. Thus they infer that they stumbled upon this same genetic feature totally independently.
Inference from a Paradigm
Of course when they say “evolution acted on” all they mean is they found the same genetic trait in two distantly related species. The notion that “evolution did it” is an inference based upon their paradigm, not a deduction from hard data. The technical paper further elaborates:
Lepidoptera is a well-known example of convergent evolution, but as studies are restricted to a few closely related species, it is difficult to make general inferences about the predictability of evolution in this system. Here we study convergent evolution in multiple mimetic neotropical lepidopteran lineages that diverged between ~1 and 120 Mya, including seven species of Ithomiini and Heliconius butterflies and a day-flying Chetone moth. Across butterfly lineages that diverged up to ~30 Mya, the genetic variants most strongly associated with convergent color pattern switches are located in similar noncoding regions near the genes ivory and optix. In the more distantly related moth species, color pattern variation is associated with a ~1 Mb inversion which also contains ivory, closely mirroring the supergene architecture of the co-mimetic butterfly Heliconius numata.
Inheritance from a Common Ancestor?
How are we to explain this evidence — genetic traits shared by distantly related insect species? Evolutionary biologists always prefer to explain the distribution of traits shared across different organisms via inheritance from a common ancestor. There’s a good reason for this preference: Darwinian evolution is at heart based upon random variation, and they recognize that it’s unlikely for the same trait to appear multiple times. As we’ll see below, even Richard Dawkins admits this point: “it is vanishingly improbable that exactly the same evolutionary pathway should ever be travelled twice.” This is precisely why evolutionary trees seek to minimize the number of total evolutionary events — because they don’t want to have to say that the same evolutionary change happened multiple times, independently.
But sometimes common ancestry can’t explain the distribution of traits. This is usually because shared traits are found in supposedly distantly related species where it’s unparsimonious to postulate that their supposed common ancestor had that trait, because other more closely related species don’t have the trait.
Evolutionary biologists typically ascribe convergent evolution to exposure to similar selection pressures. And that is exactly what this paper does here:
Convergent or parallel evolution is a natural experiment where unrelated species independently evolve similar traits in response to similar selective pressures. It informs us about the extent to which evolution is repeatable and thus predictable. Highly divergent lineages can show strong trait convergence, for example, associated with the repeated colonization of land, water, or air or the repeated evolution of resistance to challenges like insecticides or drought and heat stress.
“Vanishingly Improbable”
This sounds plausible on its face — but again, you have to understand that there’s a reason why evolutionary biologists always prefer to explain shared traits through common ancestry: convergent evolution is unlikely. Richard Dawkins explained this when he wrote in The Blind Watchmaker:
it is vanishingly improbable that exactly the same evolutionary pathway should ever be travelled twice. And it would seem similarly improbable, for the same statistical reasons, that two lines of evolution should converge on exactly the same endpoint from different starting points
It is all the more striking a testimony to the power of natural selection, therefore, that numerous examples can be found in real nature, in which independent lines of evolution appear to have converged, from very different starting points, on what looks very like the same endpoint.
Despite the improbability, both the paper and Dawkins attribute the convergent similarities to natural selection. But this is not their preferred explanation because, again, as Dawkins notes, “it is vanishingly improbable that exactly the same evolutionary pathway should ever be travelled twice,” and yet he says that “numerous examples can be found in real nature.” This is why Conway Morris says convergence is “ubiquitous” — and he even says that it has other implications:
I believe the topic of convergence is important for two main reasons. One is widely acknowledged, if as often subject to procrustean procedures of accommodation. It concerns phylogeny, with the obvious circularity of two questions: do we trust our phylogeny and thereby define convergence (which everyone does), or do we trust our characters to be convergent (for whatever reason) and define our phylogeny? As phylogeny depends on characters, the two questions are inseparable. … Even so, no phylogeny is free of its convergences, and it is often the case that a biologist believes a phylogeny because in his or her view certain convergences would be too incredible to be true. …
During my time in the libraries I have been particularly struck by the adjectives that accompany descriptions of evolutionary convergence. Words like, ‘remarkable’, ‘striking’, ‘extraordinary’, or even ‘astonishing’ and ‘uncanny’ are common place…the frequency of adjectival surprise associated with descriptions of convergence suggests there is almost a feeling of unease in these similarities. Indeed, I strongly suspect that some of these biologists sense the ghost of teleology looking over their shoulders.
Simon Conway Morris, Life’s Solution: Inevitable Humans in a Lonely Universe, pp. 127-128 (Cambridge University Press, 2003)
Two Crucial Implications
So convergent evolution has two crucial implications for evolutionary biology.
First, it implies that often shared similarities don’t imply necessarily common ancestry. Yet all evolutionary trees assume (at first, at least) that shared similarities DO indicate inheritance from a common ancestor. If there are so many exceptions to the basic assumption used to build evolutionary trees, can we really trust those trees? Convergent evolution fundamentally undermines the logic used to build trees and infer evolutionary relationships.
Second, Conway Morris (who again, does not support intelligent design) proposes that similarities shared by distantly related structures suggest some kind of teleology or purpose in nature. But we ID proponents can take it even further.
There is a known cause that can implement similarities in entirely distinct systems: intelligent agency. Intelligent agents frequently reuse similar parts in different systems. We could envision innumerable examples:
- Wheels re-used on cars and airplanes
- Power cords re-used in toasters and microwaves.
- Transistors re-used in televisions and computers
- Keyboards re-used on laptops and cell phones
- Single-piston engines being re-used in lawnmowers and motorcycles.
- Etc., etc., etc., etc., etc.
The examples of this are nearly endless. In fact, this practice of re-usage of functional parts in different systems is “ubiquitous” in human-designed technology just as convergence is “ubiquitous” in biology. We ID proponents often call such re-usage “common design.” Unfortunately, as the PLOS Biology paper shows, this option is rarely considered:
Trait convergence in different species can be caused by genetic changes at different genes or the same gene (“gene reuse”). Gene reuse is predicted to be more common among closely related lineages or when developmental pathways towards shared fitness optima are constrained. Where genes are reused, convergence may result from independent mutations at the same gene or because the same alleles are reused (“allele sharing”), either from ancestral standing variation, or as a result of introgression between species.
In other words, they attribute these shared similarities to (a) independent mutations, (b) inheritance from a common ancestor, (c) interbreeding. But they don’t consider the possibility of common design.
In the table below I explain one way of comparing the explanatory power of common design, common ancestry, and independent mutations.
In the table below I explain one way of comparing the explanatory power of common design, common ancestry, and independent mutations.
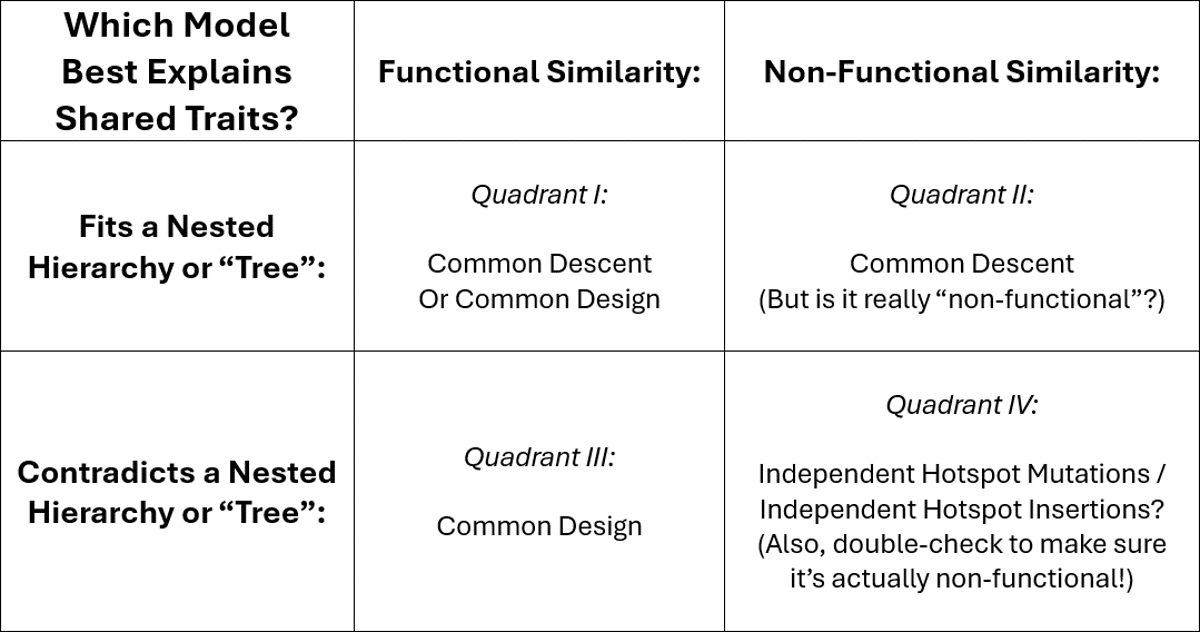
Of course, the example discussed in the paper we’re covering falls in Quadrant III — where common design is the best explanation. Much work remains to be done to flesh out a theory of common design, but the fundamentals are there and it has much promise to explain data points that evolutionary biology otherwise finds are “vanishingly improbable.”