







































Human embryonic development requires regulated activation of the zygote’s genome through chemical and mechanical inputs, establishing the signaling patterns that guide cells into forming all of the fully functional organ systems. It is these same types of mechanisms that are also responsible for the development of the nervous system.
A recent article at Science Daily, reporting on research findings in Nature Materials, demonstrates that the “force-sensitive protein called Piezo1…helps to maintain the structure of brain tissue…but also…reveals a powerful link between the brain’s physical environment and how its wiring is built.”
As I detailed in my last article here (on childbirth) about the presence of Piezo1 within uterine muscle,
Piezo1 is a large (more than 2,500 amino acids) three-bladed, propeller-shaped transmembrane protein with a central (ionic) pore at the hub. Activation of the molecular propellers, by mechanical tension on the cell membrane, opens the central pore to let calcium ions flood into the cell. This leads to downstream signaling pathways that cause a change in function.
The Science Daily article confirms that, on detecting tissue stiffness, Piezo1 ion channels within developing neural tissue “can trigger the production of guidance molecules.” This explains the underlying mechanism that scientists consider to be “a hidden force that helps wire the brain.”
This discovery not only invites examination of how this works in practical terms, but also raises the deeper question of how such an integrated system could have originated in the first place.
Growing up Neuron
The Science Daily article states:
As the brain develops, neurons grow long extensions known as axons. These structures connect different regions of the brain and transmit signals both within the brain and throughout the body. To establish these connections, axons must travel along specific routes through brain tissue.
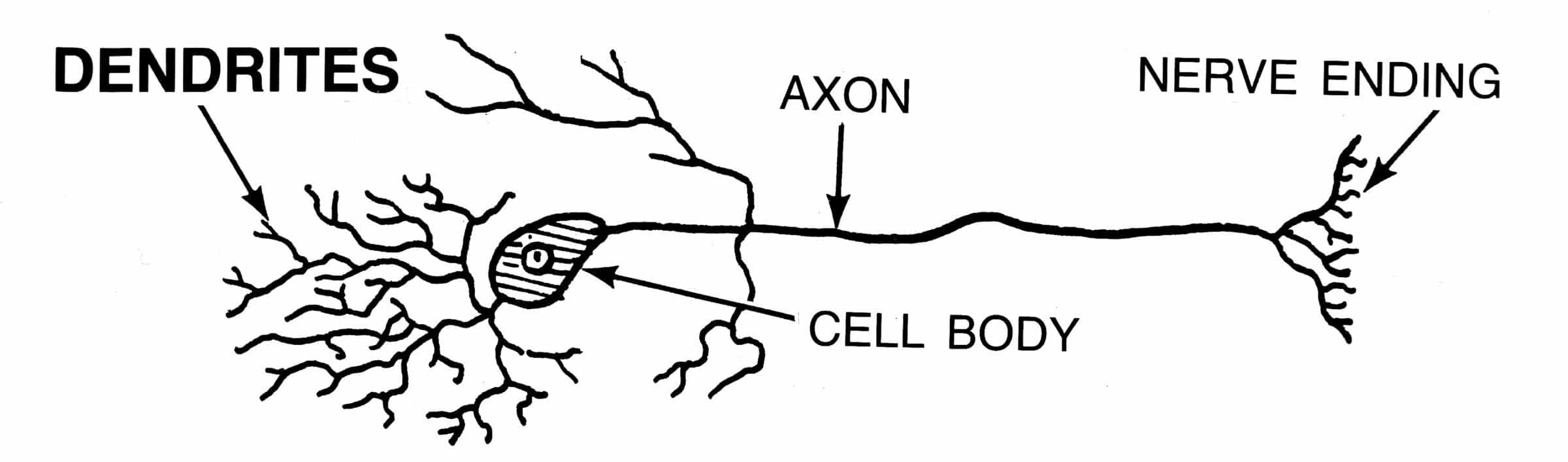
For example, as noted in the Nature Materials article, the axons of specific retinal cells in the frog exit the back of the eye and move toward the brain. Since their eyes are located on each side of their head (not in the front like ours), the axons converge backwards towards the midline. They eventually cross over to the other side of the brain and then gradually turn outward and upward to connect with the midbrain (optic tectum) where visual signals are processed and applied.
But how do axons find the right path to connect with where they are supposed to go?
Axon Trailblazing
The axon growth cone is the actively mobile, sensory navigation aid at the tip of the developing axon that ekes out a path through the extracellular matrix (ECM) to its target. It has receptors to detect guidance molecules that either attract or repel the axon, and other signaling chemicals, sent out by the surrounding neural tissue. The axon growth cone can also sense the degree of local ECM stiffness, cell tension generated by the cytoskeleton, and contact cues from nearby cells and the ECM.
The axon growth cone continuously analyzes and integrates this sensory information which ultimately signals it to continue forward, stop, or change direction (e.g., turn, veer, or retract). This requires coordinated changes within its movement machinery (cytoskeleton) and grip systems (adhesion factor function).
When an axon growth cone reaches its intended target (e.g., a nerve, muscle, or gland cell), it detects a specific biochemical “barcode” on the target cell’s membrane, along with matching molecular cues in the surrounding ECM. This combination of signals tells the growth cone that it has arrived at the right spot. In response, the axon growth cone and the cell membrane of the target cell at that site remodel their structures to form a synapse — a specialized junction with a tiny gap that allows the neuron to send chemical signals that control the target cell’s activity.
So, what controls the various guidance molecules and other mechanical factors for this to work?
Enter Piezo1
As Science Daily observes:
For many years, scientists have known that chemical signals guide how tissues grow and organize. Gradients of signaling molecules act like directional cues, helping cells move and develop in the correct locations. More recent studies have shown that physical factors such as tissue stiffness, also influence how cells behave. However, the relationship between these mechanical cues and chemical signals has remained unclear. Researchers investigated this question. Their experiments showed that tissue stiffness can regulate the production of important chemical guidance cues. This process is controlled by a mechanosensitive protein called Piezo1.
Well, how does Piezo1 accomplish these newly discovered functions?
Piezo1 —> Semaphorin-3A, Neural Cell Adhesion Molecule (NCAM1), and N-cadherin
Semaphorin-3A is a secreted glycoprotein (771 amino acids) that regulates immune function, blood vessel development (angiogenesis), skeletal biology, and axon guidance. When it binds to its receptor complex it activates intracellular signaling pathways that reorganize the cytoskeleton within the axon. This ultimately causes collapse of the growth cone which steers the axon away from regions that have high levels of Semaphorin-3A. This is why it’s called a “chemorepellent.”
NCAM1 (858 amino acids) and N‑cadherin (906 amino acids) are transmembrane glycoproteins that work as “molecular Velcro” for cell–cell adhesion within muscle, kidney, and neural tissue.
High surface levels create strong, stable binding that lets the axon growth cone grip the ECM surface and move forward. Low surface levels weaken binding, reducing traction so the axon growth cone can turn, veer, or retract.
Science Daily reports that according to the recent study, by Piezo1 sensing neural tissue stiffness, it directly impacts the concentration of guidance molecules, like Semaphorin-3A, and adhesion molecules, like NCAM1 and N-cadherin within the ECM. This ultimately affects tissue stiffness and how the axon growth cone reaches its target:
The results indicate that Piezo1 performs two important roles. It acts as a sensor that converts mechanical signals from the surrounding environment into cellular responses. At the same time, it functions as a modulator that helps organize the mechanical properties of the tissue itself.
Surprise, Surprise, Surprise!
The three members of the research team experienced awe and wonder on discovering how Piezo1 affects neural tissue and its wiring. That’s evident from their comments in Science Daily (my emphasis):
- “We didn’t expect Piezo1 to act as both a force sensor and a sculptor of the chemical landscape in the brain.”
- “What’s exciting is that Piezo1 doesn’t just help neurons sense their environment — it helps build it.”
- “This study may lead to a paradigm shift in how we think about chemical signals, with implications for many processes from early embryonic development to regeneration and disease.”
Their amazement certainly seems justified when you consider not only how an individual axon finds its way to its target cell, but how this happens throughout the entire nervous system, as reported by Stuart Burgess in his recent book Ultimate Engineering (pp. 207, 209):
One challenge with any complex mechanical system is to get wiring to every motor and sensor. In the human body, this task is unimaginably great. The human body has nerve pathways from the brain to every cubic millimeter of the body. For the nervous system to be so finely branched that it can reach every cubic millimeter of the body is astounding. It is impossible to comprehend the level of wiring detail. But to give some idea of its complexity, consider that a Boeing 747 jet has around 170 miles of wiring. That is a lot of wiring, but it’s dwarfed by the wiring in the human body, which has some 90,000 miles of wiring. Laid out end to end, that would stretch more than three times around the earth.
When one experiences amazement like that on encountering something that is more complex than expected, it usually engenders a certain amount of child-like curiosity and humility. This can often give rise to a readiness to seriously reevaluate one’s prior assumptions and a willingness to reconsider new models of causation. The whole enterprise is being embarked on to unconsciously satisfy the underlying emotion that started it in the first place. That is what usually sparks investigation.
Unfortunately, for Darwinists, expressing this emotion is usually as far as they can go along the path. For with each new discovery that confirms to scientists that life is more complicated than previously thought, the new information has to somehow be incorporated into the Darwinist origin narrative.
Darwinian evolutionary development (evo‑devo) generally proposes that axon pathfinding arose through the co‑option (repurposing) of pre‑existing molecules, gene duplication followed by changes in function, selection for advantageous variants, and the gradual assembly of large‑scale molecular networks over long periods of time. It also assumes a distributed control system in which multiple inputs (e.g., DNA, gene regulatory networks, tissue geometry, chemical gradients, and cell behaviors) just happened to get it right, again and again and again, not only for axon pathfinding, but for all of neural tissue development and for the body’s fully functioning organ systems.
A Descriptive Narrative
When scrutinized closely, the evo-devo “explanation” just uses reverse engineering to figure out how it works (even though there is still a lot that is unknown) and then provides a descriptive narrative of possible origins. It does this without any attempt to explain where the information came from to make all the parts just right, how the correct assembly took place, how the axon pathway and recognition at the target cell came about, nor how this could all evolve gradually.
This is why Darwinists can easily incorporate new discoveries into their origin narrative. It is because they never really address the true causal implications. And that’s why their emotional response to great discoveries, like this one, can only take them so far towards what seems to be the obvious.
Darwinists will tell you that Piezo1, Semaphorin-3A, NCAM1, and N-cadherin, along with their specific receptors and associated signaling patterns, all emerged through selective pressure and were then conserved over time. But this does not address how each of these parts fits into an interdependent coherent system to produce the finely tuned precision design logic needed for proper neural wiring, a process vital for survival.
In his book Undeniable (p. 20), Douglas Axe describes the “Universal Design Intuition.” As he writes, “Tasks that we would need knowledge to accomplish can be accomplished only by someone who has that knowledge. In other words, whenever we think we would be unable to achieve a particular useful result without first learning how, we judge that result to be unattainable by accident.”
That’s wisdom to live by!