Discovery Institute senior fellow Jay Richards has an excellent piece at The American titled, “When to Doubt a Scientific ‘Consensus’,” that gives 12 criteria to help us decide whether it’s appropriate to doubt a particular “consensus.” Richards of course notes that the very term “consensus” is often used to shut down scientific debate–but that hardly means the scientific “consensus” is necessarily wrong. Indeed, some wrongly challenge the consensus when it ought to be affirmed. Richards threads this needle carefully, explaining why we must carefully examine the scientific, sociological, rhetorical, and political dynamics of a debate to determine if the consensus deserves our assent, or our skepticism: Anyone who has studied the history of science knows that scientists are not immune Read More ›
Remember the analogy of the two moons I used yesterday to discuss the distribution of SINEs in the mouse and rat genomes? Well, I am going to use it again today, but only for a moment.
Moon Mysteries and the Lunarlogos Foundation
Suppose you are keenly interested in the topography of one of the moons, named Y6-9. Suppose also that the books you first select to read on the topic are popular works, written by “experts” who are “living legends.” As you read through the works, you find paragraphs here and there about how utterly decrepit Y6-9 is, and how this space body exemplifies eons of random events. The authors argue that we already knew all there was to know about that moon back in 1859, and that the evidence demonstrates either that God doesn’t exist or that the deity left the cosmos to itself after the Big Bang. You find, however, that these books almost totally ignore the findings of the billion-dollar missions sent to the surface of Y6-9 since the 1960s. Indeed, there is next to nothing in them about Y6-9’s actual geology.
So you contact the Lunarlogos Foundation, a Christian group that promotes such books. You tell them that you have a few specific questions about the Y6-9 mission findings. The response you get is that because you are a layman, you would not be able to comprehend the details. Besides, the Lunarlogos folks say, the mainstream experts have spoken authoritatively about the subject and that should be enough for you. As a consolation, though, they send you a CD that has songs that are sung by one of their founding members.
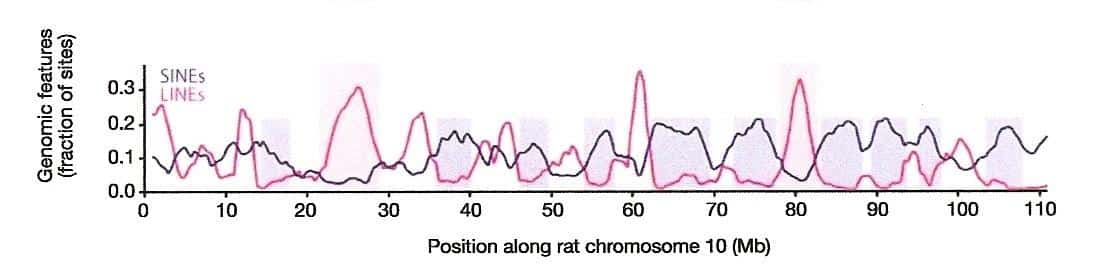
Yesterday I promised that I would show you a mysterious genomic signal, and today I shall fulfill that promise. The previous blog was devoted to describing the linear distribution of LINEs and SINEs along mammalian chromosomal DNA. We saw that L1 retrotransposons tend to be densest in the regions where Alus and Alu-like elements are the least common and vice versa. I included the following figure from an article co-authored by Francis Collins1 that showed this compartmentalization of LINEs and SINEs along over a hundred million genetic letters of rat chromosome 10:
The blue line indicates the distribution of SINEs along a 110-million base pair interval of rat chromosome 10. (From Fig. 9d of Ref. 1.)
Taxon-Specific Elements: The SINEs Aren’t The Same
Intriguing as this non-random distribution of repetitive elements may be, it gets even more interesting when one realizes that SINEs are specific to taxonomic groups. Each primate genome has distinct subfamilies of the Alu sequence. The mouse genome, on the other hand, has no Alus but it does have three unique SINE families called B1, B2, and B4. While mouse B1 shares some sequence similarity with Alu, it has no relationship to the B2 or B4 elements; the latter two are also unrelated to each other. What then about the rat SINEs along chromosome 10, which were depicted as a blue line? Well, the genome of the rat has one main SINE family called ID, for the “Identifier” sequence. The ID elements have nothing in common at the DNA sequence level with the mouse B1s, B2s, or B4s, and they are wholly dissimilar to Alus.
Brain Cancer Chromosomes. Chromosomes prepared from a malignant glioblastoma visualized by spectral karyotyping (SKY) reveal an enormous degree of chromosomal instability -- a hallmark of cancer. Created by Thomas Ried, 2014
In his recent response to Stephen Meyer’s Signature in the Cell, Francisco Ayala claimed that repetitive portions of our DNA called “Alu” sequences are “nonsensical.” Ayala wrote: “Would a function ever be found for these one million nearly identical Alu sequences? It seems most unlikely.” In his response to Ayala, Meyer showed that Ayala is factually wrong about this. According to recent technical papers in genomics, Alu sequences perform multiple functions.
In a rejoinder to Meyer, Darrel Falk defended Ayala and claimed although “a number of functional regions have been discovered within Alu sequences,” there “is no question that many Alu sequences really have no function.”
In my last blog, I showed that the vast majority of the genome is transcribed, either into protein-coding genes or into regulatory RNAs. The technical literature — some of which I cited in that blog — reports that the genome is an RNA-coding machine. Clearly, most DNA really does have function.
In this and subsequent posts, I will provide other sorts of evidence that so-called “junk DNA” is not junk at all, but functional.
We have all seen a variant of the plot in a movie. A strange signal appears–in one film it is a recurrent wireless telegraph code that is transmitted from San Diego after a global nuclear holocaust (On the Beach); in another it is radio transmissions from deep space (Contact); in still another it is crop circles (Signs). As we all know, the first signal turns out to be due to a Coca-Cola bottle: Wind blowing on a window shade next to the bottle results in the latter being occasionally nudged, which sometimes leads to a telegraph key being tapped by the very same. But in the second movie, the signals received turn out to contain a complex set of encrypted data with an intricate mathematical pattern — they are the specifications for building a device that can travel through space-time wormholes, sent from a friendly alien civilization. So also are the crop circles in the third film messages from an extraterrestrial race, except that the designs portend an attack on humanity.
Now, the reason we are drawn in by such stories is obvious: The signals have serious implications for the characters. It could mean the survival of mankind after a thermonuclear war; it could mean that there are other sentient beings in the universe. That is why we would quickly lose interest in the plot if, say, in every scene where a scientist appeared before an important governmental group and said, “The outer space signal contains over sixty thousand, multidimensional pages of complex architectural plans,” she were countered with, “This is exactly the predicted outcome of billions of years of cosmic evolution — you see, random interstellar events lead to just this kind of complex specified information…we are not impressed.” We would want our money back.
My purpose for bringing up this subject is that I have a mysterious genomic signal for you to see — which I will show you tomorrow. We detected it some time ago and it has aroused the interest of some genomicists, but you will find no mention of it books such as Francis Collins’s The Language of God — which is peculiar. But I have another aim in mind, too, for broaching this possible chromosomal code: A key first indicator of functionality is a distinctly non-random pattern. The persistence of a distinct signal in different contexts often suggests functional constraints are operative–that is why genomicists look for them. And since I want to focus on the global functions of such Short Interspersed Nuclear Elements (SINEs) as human Alus and their mouse and rat counterparts, their far-from-random placement cannot be elided. In fact, I will argue that it is a critical part of the genome story that the folks at Biologos aren’t telling you.
To prepare for the mysterious genomic signal, though, I want to draw your attention to this figure:
What you are seeing are the relative densities of Long Interspersed Nuclear Element (LINE) L1s and SINEs along 110,000,000 DNA letters of rat chromosome 10.1 (From Fig. 9d of reference 1.) The x-axis represents the sequence of letters in DNA and the blue line indicates where SINEs occur — what Ayala calls “obnoxious sequences” that are supposedly due to “degenerative biological processes that are not the result of ID.” The red line indicates where LINE sequences occur.
By the way, Francis Collins is a principal author of the Nature paper where these results are published.
Both LINEs and SINEs are types of mobile DNA, namely, retrotransposons, and together they can comprise around half of the mammalian genome. As should be clear from the figure, LINEs tend to peak in abundance where SINEs taper off and vice versa (see the blue boxes). We have known about this pattern since the late 1980s, so it is no surprise to someone who has been following the subject. What should be surprising to anyone, however, is that the same machinery is responsible for the movement of both types of retrotransposon. A complete L1 element encodes the proteins necessary to “reverse transcribe” an RNA copy of itself back into DNA, and to insert the generated duplicate into some chromosomal site. SINEs, by way of contrast, rely on the L1-specified proteins for all their copying and pasting routines.
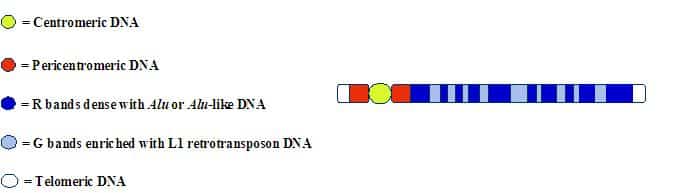
This compartmentalization of LINEs and SINEs along the mammalian chromosome can also be detected by using molecular probes for L1 or Alu(-like) sequences2:
For junkety-junk elements that can make up fifty percent of a mammal’s mostly junkety-junk genome, the rule seems to be: Location, location, location.
Interestingly, this higher-order pattern cannot be detected when small sections of DNA are examined. It only becomes evident when stretches that are millions of nucleotides long are studied. This banding pattern has been known for decades–but for some reason it is rarely (if ever) discussed by “junk DNA” advocates. The bands on the chromosome arms fall into two general categories:
A biology educator recently wrote me asking how we test intelligent design using the scientific method, how ID is falsifiable, and how ID explains patterns we observe in nature. These are very common questions that we receive all the time from teachers, students, and interested members of the public, and they’re usually legitimate, sincere, and thoughtful questions. In this case, they certainly appeared to be such, and below I post a slightly modified version of my response to the teacher, withholding any information about the teacher to protect his/her identity: We help many educators to better understand the debate over evolution. Contact us for more information!
I have long questioned the assumption that most genomic DNA sequences are “nonsensical” or “junk.” And given the data that have emerged over the past seven or so years, a functionalist view of genome has robust empirical support. It is for this reason that I think many of the arguments presented by the Biologos Foundation are “wrong on many counts,” to borrow a phrase from Darrel Falk.
Here is an example. While reading the “critique” of Steve Meyer’s book, Signature in the Cell, by Francisco Ayala, a number struck me that I know to be incorrect. The integer that I am referring to is “25,000” and it is claimed to be the known tally of genes in our chromosomes:
The human genome includes about twenty-five thousand genes and lots of other (mostly short) switch sequences…
Now, the problem with such a statement is this: While there are ~25,000 protein-coding genes in our DNA, the number of RNA-coding genes is predicted to be much higher, >450,000.1 Some of the latter range in length from being quite short–only 20 or so genetic letters–to being millions of letters long. Since 2004 we have learned that over 90% of our DNA is transcribed into RNA sequences at some developmental stage, in different cell and tissue types.2,3,4 (Our brain cells are unusually rich in these non-translated RNAs.) These RNAs are then processed into regulatory and structural sequences of all sizes.2,3,4
For over a decade, mathematician Jeffrey Shallit has been an outspoken critic of intelligent design. Recently, in a series of blog posts, he has attacked Stephen Meyer’s book Signature in the Cell (SITC) for what he sees as a variety of shortcomings. Some of Shallit’s criticisms merit careful attention, which we’ll present here in weeks to come.
Other criticisms, however, are fluffy confections, failing to achieve even the slightness of what Hume called “mere cavils and sophisms.” Let’s look at one such bonbon of sophistry, Shallit’s claim that weather forecasting represents a devastating counterexample to SITC’s argument that complex specified information is, universally in human experience, produced by a mind or intelligence.
I’ve followed the back and forth between Francisco Ayala and Steve Meyer with interest. I happened to have just read Meyer’s book Signature in the Cell when I first saw Ayala’s commentary/review on it at the Biologos Foundation website. My initial response was that Ayala obviously hadn’t read the book, and, as a result, made some embarrassing mistakes that any reader of the book would recognize.
Darrell Falk at the Biologos Foundation was apparently responsible for inviting Ayala to comment on Meyer’s book, and has been drawn into the debate.
He published the first part of Meyer’s response to Ayala, but not without first offering his “background comments” about the debate. (I think David Klinghoffer has said what needs to be said about that.) The Biologos Foundation is committed to the “science-and-religion dialogue.” In my opinion, however, they have a peculiar way of fostering dialogue.
Earlier this week, the Biologos Foundation posted part of Stephen Meyer’s response to a review of his book Signature in the Cell by evolutionary biologist Francisco Ayala. Because Biologos decided to introduce its partial posting of Meyer’s response with a misleading and inaccurate preface, and because Biologos further decided to rebut part 1 of Meyer’s response before readers had a chance to read his entire response, we have decided to make the rest of Dr. Meyer’s response available on his website immediately. Just as readers were allowed to read Dr. Ayala’s critique in its entirety before reading Dr. Meyer’s response, we think it only fair that readers should have the opportunity to read Meyer’s entire response (which was written in Read More ›
Discovery Institute has invited Dr. Francisco Ayala to debate the thesis of the book Signature in the Cell: DNA and the Evidence for Intelligent Design with the book’s author, Dr. Stephen Meyer. Those who’ve been following the debate between Meyer and his critics know that there has been a bit of back and forth since Ayala was invited to critique SITC on the Biologos website. Meyer has responded this week, with the first of two parts on the Biologos site. Discovery Institute would like to initiate a full-fledged, official debate between the two, and so we have already sent the following invitation to Dr. Ayala. Dear Professor Ayala: I am writing to you in my capacity as the Director of Read More ›