Francisco J. Ayala, biologist and former Dominican priest, has won this year’s Templeton Prize. Valued at $1.53 million, the prize has sought to reward serious thought, writing and research pointing the way to a reconciliation of science and faith. In Ayala’s case, for “science” read “Darwinism.” So a word or two is in order about the faith of Dr. Ayala.
Advocates of a supposedly religion-friendly Darwinism have seized on the idea of God’s acting through secondary causes. In his book Darwin’s Gift to Science and Religion, Ayala argues that since God acts through intermediate causation to create geological features (mountains, rivers), why may the same analysis not be applied to the evolution of life? In the latter context, he insists that the idea of God’s acting through “specific agency…amounts to blasphemy.” For such direct control would imply that God bears responsibility for all the cruelties, pains, and dysfunctions that have accompanied the unfolding of life’s history.
But there is a real and important difference between secondary causation of the kind that results in the formation of rivers and mountains, on one hand, and that which, according to the evolutionary model, results in life in all its forms. The operation of geological forces follows paths described by physical laws. Whatever role chance plays, the overall process is predictable. The religious believer may reasonably picture God, having authored those laws, as the creator of geological features, having planned and foreseen what those features would be. Similarly, He is the author of those laws that govern patterns in the weather, in the alternation of the seasons, of day and night, and so on. God could thus confidently tell Noah that “So long as the earth exists, seedtime and harvest, cold and heat, summer and winter, and day and night shall not cease” (Genesis 8:6).
But life — including human life — is different. If Darwin and the vast majority of his modern advocates are right, then the path of life’s evolution was inherently unpredictable — not wholly random, since natural selection plays its role, but generated by chance and governed by no plan, design, or teleology. Ayala himself has said this very clearly: “It was Darwin’s greatest accomplishment to show that the complex organization and functionality of living beings can be explained as the result of a natural process — natural selection — without any need to resort to a Creator or other external agent.”
The failure of Darwinism to account for our human experience is something many people know intuitively — but few can articulate it so well as Harvard philosopher Harvey Mansfield and novelist Tom Wolfe. Peter Lawler, who blogs over at First Thing’s Postmodern Conservative, wrote a wonderful essay detailing the ways “America’s two most astute social commentators… have weighed in on the debate over the neo-Darwinian view of evolution.” In “Real Men Prove Darwin Wrong (Again),” Lawler synthesizes how these two masters illustrate that there are more things in heaven and earth than can be explained by Darwin:
They agree that the real controversy in our country is not between rationalists who preach evolutionism and fundamentalists who live in Darwin-denial, but between those who still believe that evolution can account for the whole of human behavior and those who see with their own eyes that it does not. The Darwinians, they observe, cannot properly account for the natural human quality that Mansfield calls “manliness” and that Wolfe, following the sociologists, describes as each individual’s concern for his own status or ranking. The Darwinians do not recognize what genuinely distinguishes the human individual from everything else in nature, so they cannot account for such admirable phenomena as Carson Holloway’s defense of transcendent human nobility against Darwinian reductionism.
Lawler’s essay is incisive and enlightening, reflecting on the denial of manliness (that character trait that drives an individual to believe that she is someone worth championing) inherent in the Darwinian fight against individualism:
Darwinians criticize the human tendency toward championism, and they fight against both our individualism and our speciesism. Science, they think, promises to free us from the illusion that there is anything special about me or mine. It frees us from our religious tendency to think God gave us a privileged place in the nature which, in truth, treats all life forms with equal indifference. The theory of evolution, according to Wolfe, is both a denial of, and a replacement for, religion. It replaces the older “championism” with the proudly dogmatic atheism of those who style themselves special enough to know that there is nothing at all special about us.
In his book Coming to Peace with Science, Point Loma Nazarene University biology professor Darrel R. Falk makes many arguments for common descent and Darwinian evolution. Many of these arguments are evidence-based (some of this evidence is countered in a previous post), but some of his most forceful arguments are not based upon evidence. They are based upon appeals to authority. Consider the following: If you’re seeing a pattern here, it’s because Dr. Falk likes to appeal to the authority of “virtually all” scientists to make his arguments. Now to Dr. Falk’s credit, he spends a lot of time talking about the evidence, but this repeated argumentum ad “virtually all” scientists is a little troubling. Now as I wrote recently, Read More ›
Recently I was e-mailed by an individual who had read the book Coming to Peace with Science, by Darrel Falk, president of the BioLogos Foundation. This person was interested in a response to the arguments for human/ape common ancestry in Dr. Falk’s book. Not having read Dr. Falk’s book before, I wrote back that I hadn’t yet read the book but had a strong suspicion that it would argue that shared non-functional (aka “junk”) DNA between humans, apes, and other species is evidence of their common ancestry. This is an extremely common argument from theistic evolutionists–Francis Collins made it in The Language of God (and Collins wrote the foreword to Dr. Falk’s book). Of course in 2010, we’re seeing more Read More ›
Unlike the reptilian Dawkins, sinister Dennett, or smug Coyne, Michael Ruse is a prominent Darwinian atheist by whom it’s hard not to feel charmed. I have never met him but his column in the Guardian could only be written by someone who is by nature very nice, very naïve, or probably both.
Ruse considers the question of what impact belief in Darwinism should be expected to have on morality, and he answers with an “on the one, on the other hand.” Since God is dead (and Darwin killed him), there can be no objective moral ideas. Moses received nothing on Sinai. Yet, not to worry! This realization will not lead to bad behavior since our genes, inscribed by natural selection, create within us a feeling, however illusory, that moral standards really are objective. Knowing that they are subjective does not dispel the impulse to be good. We feel compelled to obey ethical dictates. God’s demise is therefore not a “significant finding”:
Now you know that morality is an illusion put in place by your genes to make you a social cooperator, what’s to stop you behaving like an ancient Roman [raping Sabine women]? Well, nothing in an objective sense. But you are still a human with your gene-based psychology working flat out to make you think you should be moral. It has been said that the truth will set you free. Don’t believe it. David Hume knew the score. It doesn’t matter how much philosophical reflection can show that your beliefs and behavior have no rational foundation, your psychology will make sure you go on living in a normal, happy manner.
Of course a Darwin apologist would have to assert something along these lines. The project of defending Darwinism mandates it. What else is one going to say: “Darwinism is both true and a spreading social poison. Get used to it”? There is no constituency for that message. Various Darwinians have offered their own miscellaneous theories as to why evolutionary belief poses no threat to decency. Robert Wright, who likewise comes across as a pleasant person, has written several upbeat books contributing his own thoughts on the subject.
Discovery Institute senior fellow Jay Richards has an excellent piece at The American titled, “When to Doubt a Scientific ‘Consensus’,” that gives 12 criteria to help us decide whether it’s appropriate to doubt a particular “consensus.” Richards of course notes that the very term “consensus” is often used to shut down scientific debate–but that hardly means the scientific “consensus” is necessarily wrong. Indeed, some wrongly challenge the consensus when it ought to be affirmed. Richards threads this needle carefully, explaining why we must carefully examine the scientific, sociological, rhetorical, and political dynamics of a debate to determine if the consensus deserves our assent, or our skepticism: Anyone who has studied the history of science knows that scientists are not immune Read More ›
Remember the analogy of the two moons I used yesterday to discuss the distribution of SINEs in the mouse and rat genomes? Well, I am going to use it again today, but only for a moment.
Moon Mysteries and the Lunarlogos Foundation
Suppose you are keenly interested in the topography of one of the moons, named Y6-9. Suppose also that the books you first select to read on the topic are popular works, written by “experts” who are “living legends.” As you read through the works, you find paragraphs here and there about how utterly decrepit Y6-9 is, and how this space body exemplifies eons of random events. The authors argue that we already knew all there was to know about that moon back in 1859, and that the evidence demonstrates either that God doesn’t exist or that the deity left the cosmos to itself after the Big Bang. You find, however, that these books almost totally ignore the findings of the billion-dollar missions sent to the surface of Y6-9 since the 1960s. Indeed, there is next to nothing in them about Y6-9’s actual geology.
So you contact the Lunarlogos Foundation, a Christian group that promotes such books. You tell them that you have a few specific questions about the Y6-9 mission findings. The response you get is that because you are a layman, you would not be able to comprehend the details. Besides, the Lunarlogos folks say, the mainstream experts have spoken authoritatively about the subject and that should be enough for you. As a consolation, though, they send you a CD that has songs that are sung by one of their founding members.
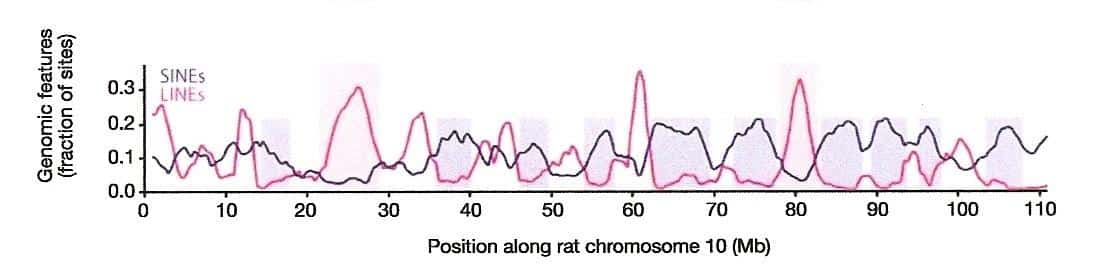
Yesterday I promised that I would show you a mysterious genomic signal, and today I shall fulfill that promise. The previous blog was devoted to describing the linear distribution of LINEs and SINEs along mammalian chromosomal DNA. We saw that L1 retrotransposons tend to be densest in the regions where Alus and Alu-like elements are the least common and vice versa. I included the following figure from an article co-authored by Francis Collins1 that showed this compartmentalization of LINEs and SINEs along over a hundred million genetic letters of rat chromosome 10:
The blue line indicates the distribution of SINEs along a 110-million base pair interval of rat chromosome 10. (From Fig. 9d of Ref. 1.)
Taxon-Specific Elements: The SINEs Aren’t The Same
Intriguing as this non-random distribution of repetitive elements may be, it gets even more interesting when one realizes that SINEs are specific to taxonomic groups. Each primate genome has distinct subfamilies of the Alu sequence. The mouse genome, on the other hand, has no Alus but it does have three unique SINE families called B1, B2, and B4. While mouse B1 shares some sequence similarity with Alu, it has no relationship to the B2 or B4 elements; the latter two are also unrelated to each other. What then about the rat SINEs along chromosome 10, which were depicted as a blue line? Well, the genome of the rat has one main SINE family called ID, for the “Identifier” sequence. The ID elements have nothing in common at the DNA sequence level with the mouse B1s, B2s, or B4s, and they are wholly dissimilar to Alus.
Despite the old canard that the only people to question Darwinian evolution are evangelical Protestants (a canard regurgitated yet again last week by the New York Times), the fact remains that Darwin dissenters can be found among thoughtful scientists and other people from all religions and walks of life. There have been many Catholic dissenters from Darwin, from St. George Jackson Mivart and G.K. Chesterton a century ago to biochemist Michael Behe and philosopher/theologian Benjamin Wiker today. There also have been numerous Jewish dissenters from Darwin. David Klinghoffer writes about one of them in an essay for First Things on Rabbi Samson Raphael Hirsch (1808—1888): Hirsch insisted again and again that God must be understood as acting with complete freedom Read More ›
Brain Cancer Chromosomes. Chromosomes prepared from a malignant glioblastoma visualized by spectral karyotyping (SKY) reveal an enormous degree of chromosomal instability -- a hallmark of cancer. Created by Thomas Ried, 2014
In his recent response to Stephen Meyer’s Signature in the Cell, Francisco Ayala claimed that repetitive portions of our DNA called “Alu” sequences are “nonsensical.” Ayala wrote: “Would a function ever be found for these one million nearly identical Alu sequences? It seems most unlikely.” In his response to Ayala, Meyer showed that Ayala is factually wrong about this. According to recent technical papers in genomics, Alu sequences perform multiple functions.
In a rejoinder to Meyer, Darrel Falk defended Ayala and claimed although “a number of functional regions have been discovered within Alu sequences,” there “is no question that many Alu sequences really have no function.”
In my last blog, I showed that the vast majority of the genome is transcribed, either into protein-coding genes or into regulatory RNAs. The technical literature — some of which I cited in that blog — reports that the genome is an RNA-coding machine. Clearly, most DNA really does have function.
In this and subsequent posts, I will provide other sorts of evidence that so-called “junk DNA” is not junk at all, but functional.
We have all seen a variant of the plot in a movie. A strange signal appears–in one film it is a recurrent wireless telegraph code that is transmitted from San Diego after a global nuclear holocaust (On the Beach); in another it is radio transmissions from deep space (Contact); in still another it is crop circles (Signs). As we all know, the first signal turns out to be due to a Coca-Cola bottle: Wind blowing on a window shade next to the bottle results in the latter being occasionally nudged, which sometimes leads to a telegraph key being tapped by the very same. But in the second movie, the signals received turn out to contain a complex set of encrypted data with an intricate mathematical pattern — they are the specifications for building a device that can travel through space-time wormholes, sent from a friendly alien civilization. So also are the crop circles in the third film messages from an extraterrestrial race, except that the designs portend an attack on humanity.
Now, the reason we are drawn in by such stories is obvious: The signals have serious implications for the characters. It could mean the survival of mankind after a thermonuclear war; it could mean that there are other sentient beings in the universe. That is why we would quickly lose interest in the plot if, say, in every scene where a scientist appeared before an important governmental group and said, “The outer space signal contains over sixty thousand, multidimensional pages of complex architectural plans,” she were countered with, “This is exactly the predicted outcome of billions of years of cosmic evolution — you see, random interstellar events lead to just this kind of complex specified information…we are not impressed.” We would want our money back.
My purpose for bringing up this subject is that I have a mysterious genomic signal for you to see — which I will show you tomorrow. We detected it some time ago and it has aroused the interest of some genomicists, but you will find no mention of it books such as Francis Collins’s The Language of God — which is peculiar. But I have another aim in mind, too, for broaching this possible chromosomal code: A key first indicator of functionality is a distinctly non-random pattern. The persistence of a distinct signal in different contexts often suggests functional constraints are operative–that is why genomicists look for them. And since I want to focus on the global functions of such Short Interspersed Nuclear Elements (SINEs) as human Alus and their mouse and rat counterparts, their far-from-random placement cannot be elided. In fact, I will argue that it is a critical part of the genome story that the folks at Biologos aren’t telling you.
To prepare for the mysterious genomic signal, though, I want to draw your attention to this figure:
What you are seeing are the relative densities of Long Interspersed Nuclear Element (LINE) L1s and SINEs along 110,000,000 DNA letters of rat chromosome 10.1 (From Fig. 9d of reference 1.) The x-axis represents the sequence of letters in DNA and the blue line indicates where SINEs occur — what Ayala calls “obnoxious sequences” that are supposedly due to “degenerative biological processes that are not the result of ID.” The red line indicates where LINE sequences occur.
By the way, Francis Collins is a principal author of the Nature paper where these results are published.
Both LINEs and SINEs are types of mobile DNA, namely, retrotransposons, and together they can comprise around half of the mammalian genome. As should be clear from the figure, LINEs tend to peak in abundance where SINEs taper off and vice versa (see the blue boxes). We have known about this pattern since the late 1980s, so it is no surprise to someone who has been following the subject. What should be surprising to anyone, however, is that the same machinery is responsible for the movement of both types of retrotransposon. A complete L1 element encodes the proteins necessary to “reverse transcribe” an RNA copy of itself back into DNA, and to insert the generated duplicate into some chromosomal site. SINEs, by way of contrast, rely on the L1-specified proteins for all their copying and pasting routines.
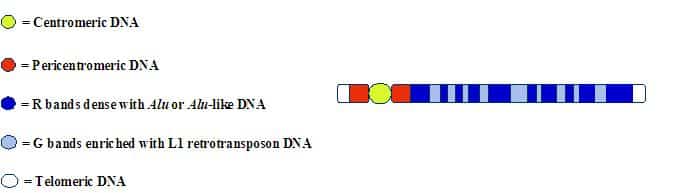
This compartmentalization of LINEs and SINEs along the mammalian chromosome can also be detected by using molecular probes for L1 or Alu(-like) sequences2:
For junkety-junk elements that can make up fifty percent of a mammal’s mostly junkety-junk genome, the rule seems to be: Location, location, location.
Interestingly, this higher-order pattern cannot be detected when small sections of DNA are examined. It only becomes evident when stretches that are millions of nucleotides long are studied. This banding pattern has been known for decades–but for some reason it is rarely (if ever) discussed by “junk DNA” advocates. The bands on the chromosome arms fall into two general categories: